



我国田间发现低致死率非洲猪瘟基因II型自然变异流行株
发布时间:
2021-03-01 14:23
来源:
维科生物
作者:
2020年,中国农业科学院哈尔滨兽医研究所国家非洲猪瘟专业实验室在开展非洲猪瘟流行病学监测及病原学研究中发现,我国部分省区出现了低致死率的非洲猪瘟基因II型自然变异流行株。这些变异株至少包括4种CD2v编码失活突变类型,导致病毒粒子失去吸附红细胞表型;相比典型强毒致病力降低,但仍具有明显的残留毒力,较高剂量接种猪可引起亚急性、慢性病程和部分死亡,较低剂量感染则主要引起持续感染和慢性病程,具有很强的水平传播能力。这些变异株临床表现具有一定的隐蔽性,早期诊断难度加大,为我国非洲猪瘟防控带来全新的挑战,必须加以高度重视,尽快制定和采取应对策略。
(相关研究论文 “Emergence and prevalence ofnaturally occurring lower virulent African swine fever viruses in domestic pigsin China in 2020” 已于2021年2月24日在线预发表在SCIENCE CHINA Life Sciences。)
非洲猪瘟(African Swine fever,ASF)是由非洲猪瘟病毒(African Swine Fever Virus,ASFV)引起的各年龄段家猪与野猪的急性、烈性、高度接触性且高发病率和死亡率的传染病,给养猪业带来严重的经济损失并威胁全球粮食安全。目前尚无商品化疫苗或有效的治疗方法。ASF被世界动物卫生组织(OIE)列为应通报疾病,世界粮食及农业组织将其列为主要跨界动物疾病。该病1914年首次报道于非洲肯尼亚,1957年从非洲传至葡萄牙,现在主要流行于非洲、东欧和亚洲,给世界范围内生猪产业带来巨大威胁。ASFV结构复杂,在环境中极其稳定。ASFV在家猪和野猪中长期流行传播,造成病毒遗传变异多样化。根据B646L基因(P72)可变3'-末端的核苷酸序列,ASFV已鉴定出至少24种基因型。目前,欧洲主要流行基因I和II型,而亚洲主要流行基因II型。
2018年8月,ASF传入我国并迅速扩散,给我国生猪产业造成了巨大的经济损失,尚无产业化应用的疫苗和药物,早期诊断及感染猪群扑杀是目前防控主要策略。流行病学监测及流行株的基因组变异、生物学表型及致病力研究对非洲猪瘟防控具有关键指导意义。中国农业科学院哈尔滨兽医研究所国家非洲猪瘟专业实验室于2018年首次分离鉴定了我国第一株非洲猪瘟病毒Pig/HLJ/2018(HLJ/18),动物回归试验证实HLJ/18感染家猪表现为急性病程及高度致死。非洲猪瘟已在我国流行了两年多,田间流行毒株的基因组变异及生物学和致病性表型进化亟待解析。
1. 非洲猪瘟病毒监测
2020年6月至12月,国家非洲猪瘟专业实验室按照农业农村部畜牧兽医局统一安排,执行《农业农村部畜牧兽医局关于调整部分省份非洲猪瘟监测工作的通知》及《农业农村部畜牧兽医局关于调整规模猪场等场所非洲猪瘟采样检测任务分工的通知》(农牧便函〔2020〕670号)的要求,对黑龙江、吉林、辽宁、内蒙古以及陕西5省进行常态化非洲猪瘟主动监测与流行病学调查,总计采集并检测、分析病原学样品3522份;同时,对部分省市送检的138份疑似阳性样品进行了鉴定及检测。所有样本使用OIE推荐的qPCR方法对病毒p72基因进行检测。ASFV DNA阳性样品利用猪原代肺泡巨噬细胞(PAM)进行病毒分离,最终成功分离到22株非洲猪瘟病毒(表1)。
2. ASFV分离株的遗传变异分析
为了分析22株病毒的遗传变异情况,选取ASFV重要的抗原基因、毒力决定基因和高突变区的23个ORF或区域进行了克隆测序。序列分析表明,所有分离株B646L基因(p72蛋白)序列与HLJ/18一致,均属于基因II型。23个ORF或区域中的15个未产生任何突变。而余下8个ORF或区域(MGF_110-14L、MGF_110-1L、EP402R、CP530R、E199L、O174L、TRS以及A238L)存在不同程度的核苷酸突变和缺失、片段插入及小片段替换情况(图1)。
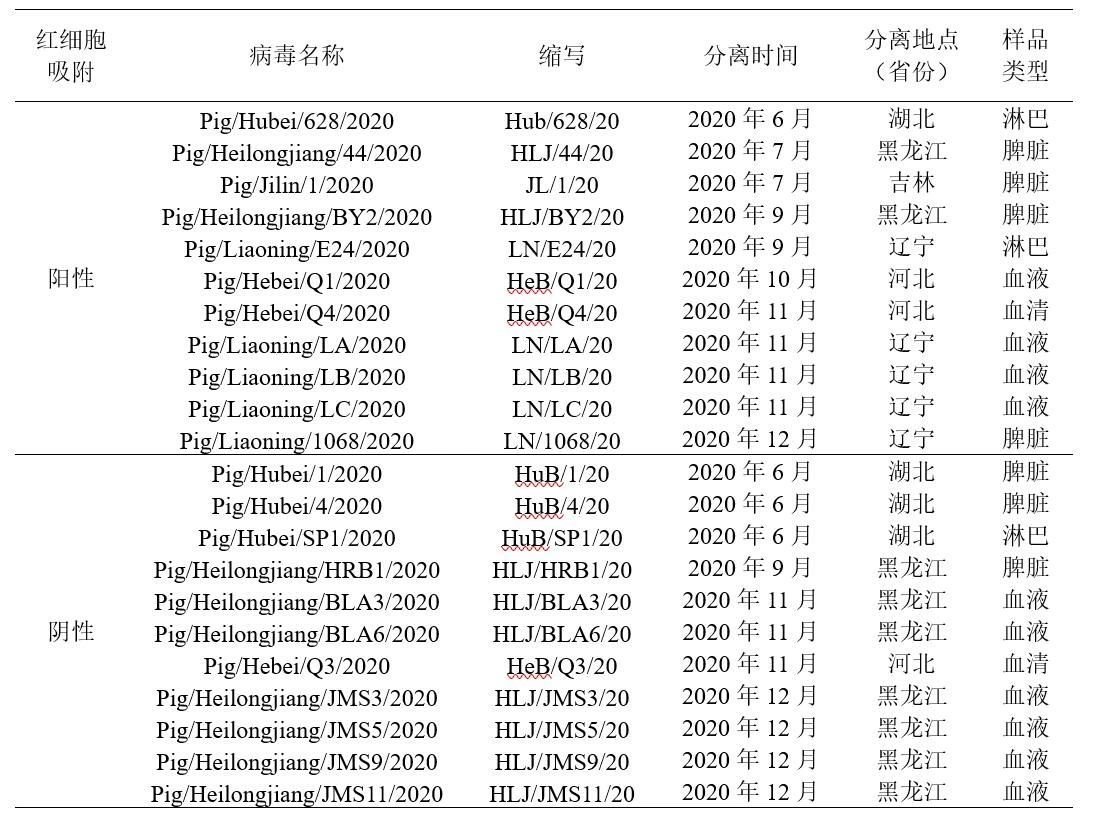

文献证明A238L基因能抑制NF-κB和NFAT介导的先天免疫反应,进而逃避宿主免疫应答。序列分析发现,HLJ/HRB1/20病毒A238L基因的536-549位发生14个核苷酸的缺失,造成其自179位氨基酸开始移码,与其它毒株相比增加了4个氨基酸(图1)。对HLJ/HRB1/20进行全基因组序列测定(GenBank 登录号MW656282),分析发现其基因组上没有明显的人工改造标记,如报告基因或标签等。
3. ASFV分离株的体外培养特性鉴定
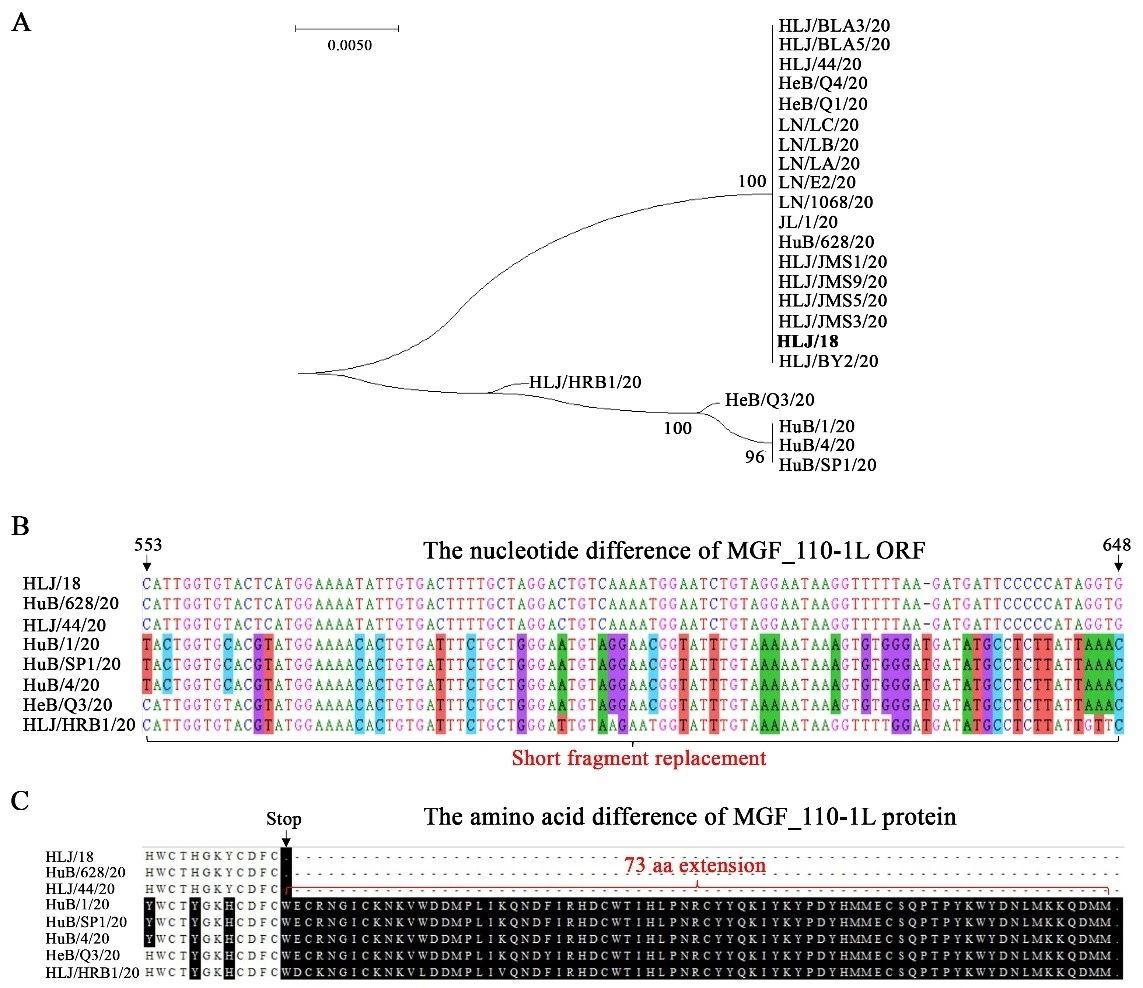
根据分离地点及基因组遗传变异特点,我们选取四株ASFV(HuB/628/20、HLJ/44/20、HLJ/HRB1/20和HeB/Q3/20)进行体外生物学特性分析。间接免疫荧光(IFA)结果显示4株病毒均可以在PAM细胞上进行有效复制(图3A)。HAD试验确定HuB/628/20和HLJ/44/20可以形成典型的HAD表型;但HeB/Q3/20和HLJ/HRB1/20呈现HAD表型阴性,不能吸附红细胞(图3A),与基因组变异分析结果一致。电镜观察四株病毒之间没有明显的形态学差异(图3A)。复制动力学显示,四株病毒可以在PAM中有效复制,感染后第五天细胞上清液中的p72基因拷贝数均大于108/mL(图3B)。

4. HAD表型阳性的分离株接种猪急性发病、高度致死
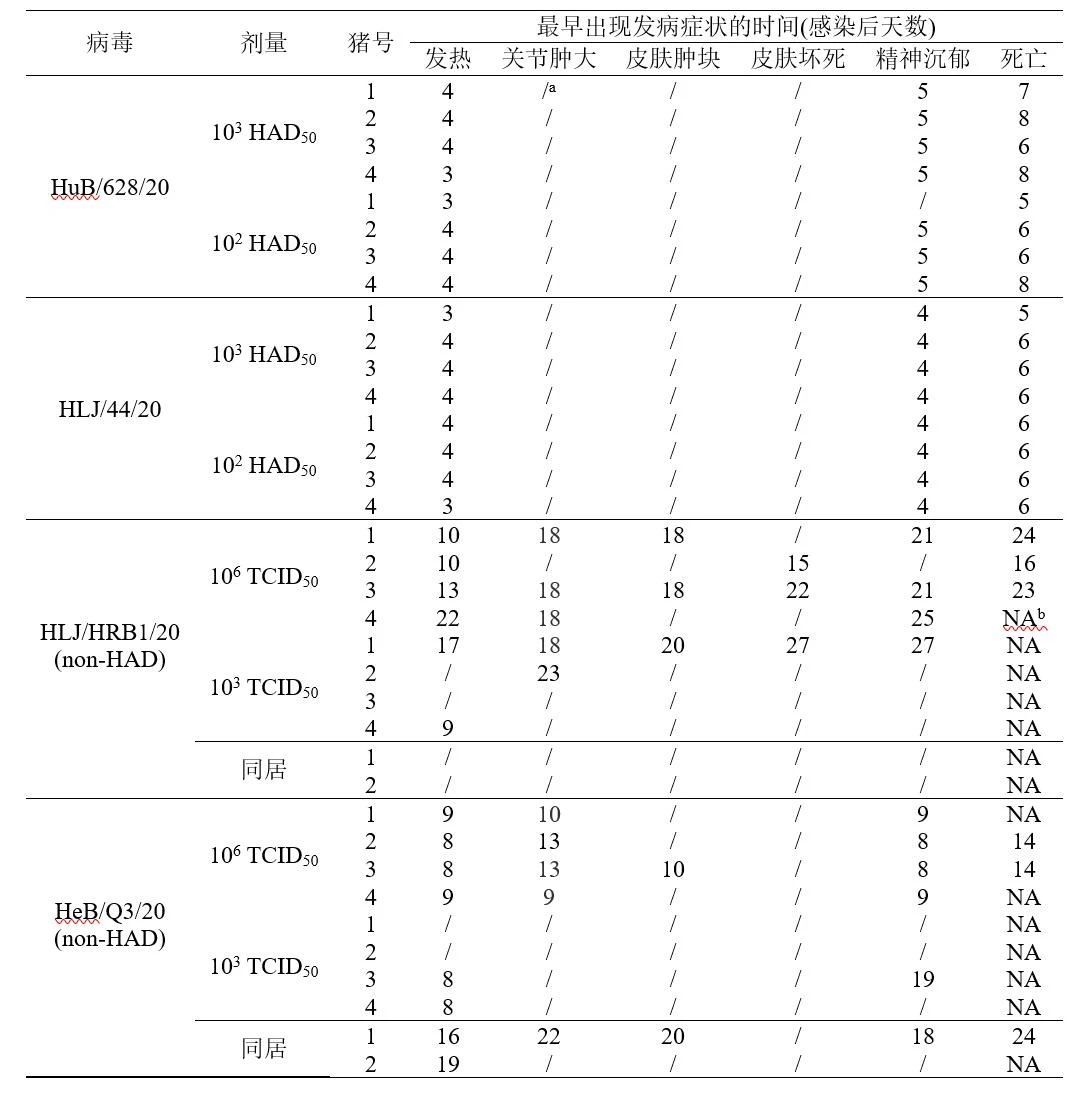
对HAD表型阳性的两株病毒HuB/628/20和HLJ/44/20,进行动物回归试验,评估其对家猪的致病力。分别以102和103 HAD50两个剂量肌肉注射接种猪,每剂量4头,每天观察接种猪临床症状和死亡情况。HuB/628/20感染猪第3天起开始出现发病症状,包括发热、精神沉郁、皮肤发绀或呼吸窘迫;感染后后第5-8天全部发病死亡。HLJ/44/20感染猪第3天起出现症状;在接种后第5-6天全部死亡(表2和图4)。结果表明,HuB/628/20和HLJ/44/20对家猪的致病力与HLJ/18相当,对家猪都表现高度致死。
5. 缺失HAD表型(non-HAD)分离株毒力降低,接种猪引起持续感染、慢性病程和部分死亡
已有研究证明CD2v是影响非洲猪瘟病毒毒力的重要功能基因之一。人工缺失CD2v编码基因可降低BA71株(基因型I)和HLJ/18株(基因型II)对家猪致病力,但CD2v编码基因缺失不影响Malawi Lil-20/1和Georgia 2010分离株的致病力。为了明确CD2v编码失活对我国田间毒株致病力的影响,我们选取2株non-HAD病毒进行动物回归试验。其中一株为CD2v编码区发生25个核苷酸缺失的黑龙江分离株HLJ/HRB1/20,另一株为CD2v基因301位突变的河北分离株HeB/Q3/20。用106TCID50和103 TCID50两个剂量分别接种SPF猪,每剂量接种4头,持续观测28天。
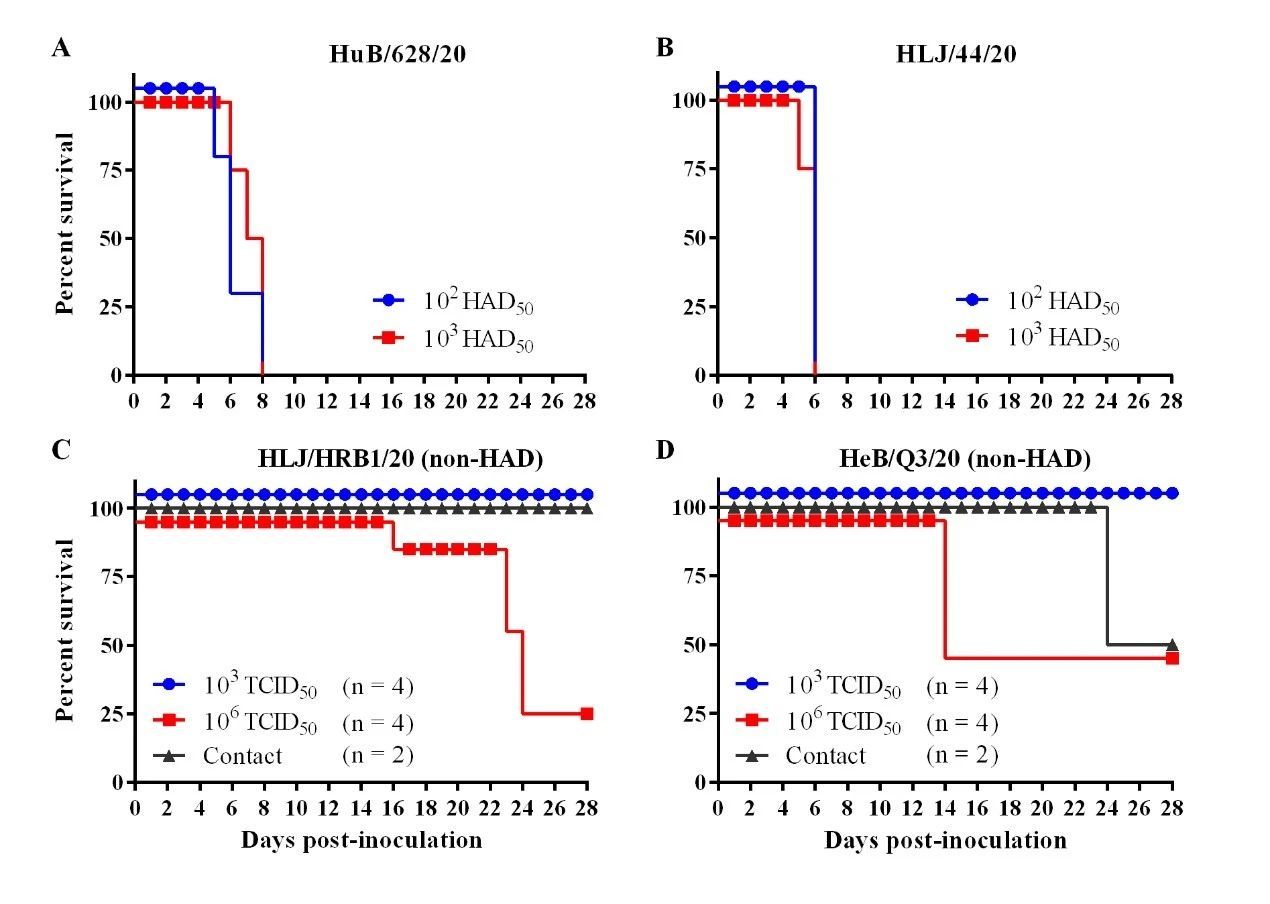
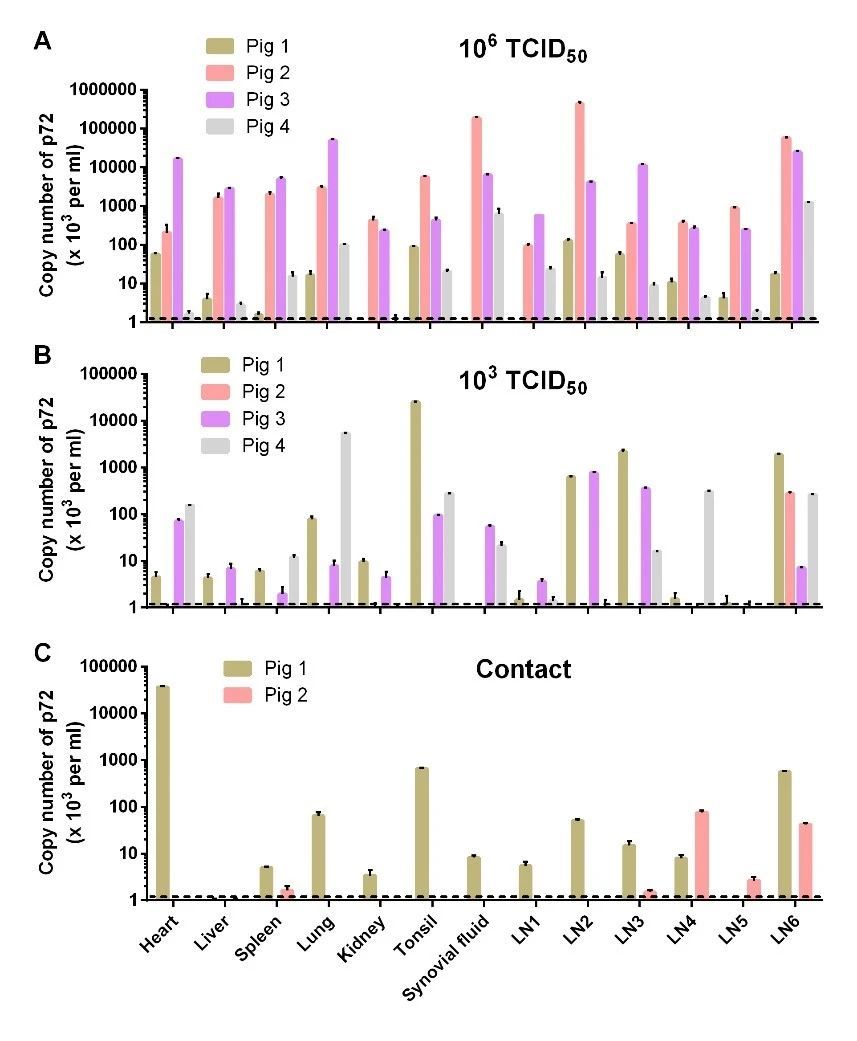
结果发现, HLJ/HRB1/20分离株106 TCID50接种组有2头猪在接种后的第10天出现持续体温升高现象(>40.5℃),第13天共有3头猪出现体温升高,另一头猪在接种后第22天出现持续体温升高。103 TCID50接种猪1头在接种后第17天出现持续体温升高,另一头猪在接种后第9-12天持续4天高温(表2)。106 TCID50 接种4头猪有3头分别在接种后第16天、23天和24天死亡,有1头猪存活;103 TCID50接种的4头猪全部存活(图4)。106 TCID50接种猪普遍出现关节肿大、腹部和胸部皮肤肿块凸起,腹部脐带处出现皮肤溃烂等亚临床症状(图5)。103 TCID50 接种猪也出现了明显的关节肿大现象。对采集的口拭子及肛拭子进行qPCR病毒核酸检测发现,106TCID50接种猪自接种后第4天开始口腔排毒,其存活猪排毒一直持续到接种后第22天;第10天开始肛门排毒,其存活猪排毒持续到接种后第24天(图6)。103 TCID50接种猪在接种后第8天开始出现口腔排毒,在接种第24天后仍有个别猪高水平口腔排毒;在接种后第10天开始肛门排毒(图6)。接种后不同时间采集抗凝血进行qPCR核酸检测,106 TCID50接种猪有2头在接种第3天血液中即可检测到低水平病毒核酸拷贝,第5天起4头猪血液中全部检测到病毒(图6)。103 TCID50接种猪血样检到病毒起始时间较高剂量组滞后2天,且血液中病毒含量低于高剂量组(图6)。
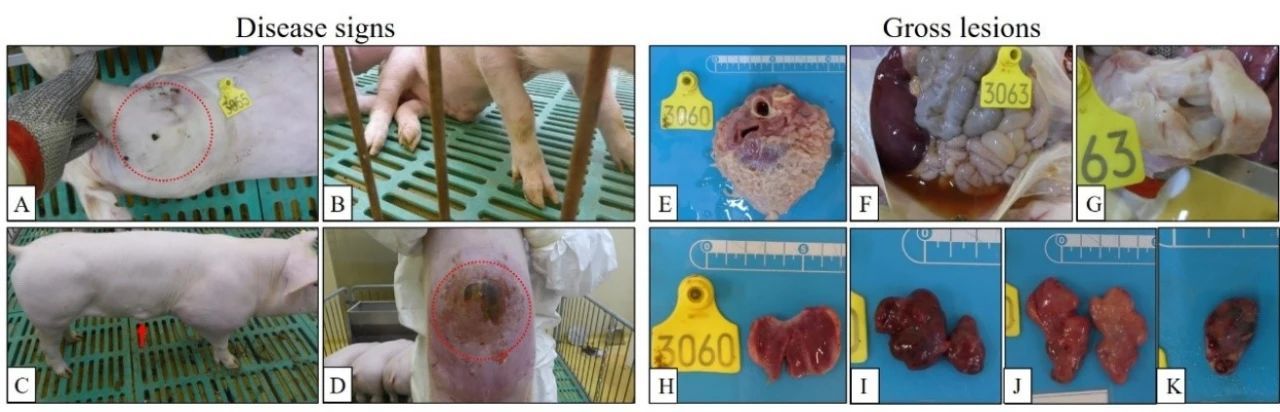
HeB/Q3/20分离株106 TCID50接种组有2头猪第8天出现持续体温升高现象,第9天4头猪全部体温升高,表现出与HLJ/HRB1/20高剂量攻毒组相似的临床症状(表2);第14天有2头接种猪发病死亡。103 TCID50接种组有2头猪第8天出现持续体温升高。103 TCID50攻毒组的4头猪28天观察期内均存活(图4);除皮肤坏死外,所有存活猪均表现出与HLJ/HRB1/20低剂量攻毒组的猪相似的病程和临床特征(表2)。
2018年分离的我国第一株非洲猪瘟流行株HLJ/18,在接种后第3天所有猪就出现体温持续升高,第4天口拭子排毒,第5天肛拭子排毒,10天之内接种猪全部死亡(Zhao D et al, EMI, 2019)。相比之下,两株CD2v功能失活自然变异株HLJ/HRB1/20和HeB/Q3/20对SPF猪致病力均显著降低。

6. HLJ/HRB1/20攻毒猪剖检病变及组织病毒载量
对HLJ/HRB1/20分离株 106 TCID50接种组的3头病死猪以及存活至28天试验猪进行剖检,组织器官病理损伤主要表现为淋巴结充血和肿胀、绒毛心、心包积液及扁桃体炎症(图5),其中2头猪还出现腹膜炎。HLJ/HRB1/20 103 TCID50接种存活猪均出现心包积液(表3)。利用qPCR对心脏、肝、脾、肺、肾、扁桃体和六个淋巴结(肠淋巴结,腹股沟淋巴结,上颌下淋巴结,支气管淋巴结,胃肝淋巴结和纵隔淋巴结)进行病毒基因组核酸检测,结果显示,在所有病死及存活猪各组织均检测到病毒核酸,106 TCID50攻毒组3只死亡猪的多种组织病毒载量明显高于存活猪;各种组织中,肺、扁桃体和部分淋巴结中检测到病毒载量相对更高(图7)。
7. 缺失HAD表型(non-HAD)分离株表现出很强接触传播能力
为了评估non-HAD病毒株接触传播能力,从感染的第一天起,分别将两头SPF仔猪与106 TCID50剂量HLJ/HRB1/20和HeB/Q3/20接种猪同居饲养,每天观察其临床体征和温度变化,采集血液、口腔拭子以及肛拭子,qPCR检测病毒核酸,评估水平传播能力。
HLJ/HRB1/20接种组的两头同居猪,分别于第6天和第12天从口腔拭子中检测到病毒核酸,第8天和第20天从肛拭子中检测到病毒核酸(图6);第21天从一头猪的血样中检测到病毒核酸(图6)。两头同居猪在同居后第28天均未出现明显的临床症状,同居后第28天进行了安乐死,剖检观察发现部分组织发生轻度病变,包括淋巴结充血、肿大、萎缩与出血点以及肺炎等(表3)。组织器官病毒载量检测发现,一头猪的心脏、脾脏、肺、肾、扁桃体、滑膜、肠道淋巴结、腹股沟淋巴结、上颌下淋巴结、支气管淋巴结、胃肝淋巴结和纵隔淋巴结均检测到病毒核酸;而另一头猪,主要在支气管淋巴结和纵隔淋巴结中检测到病毒核酸(图7)。
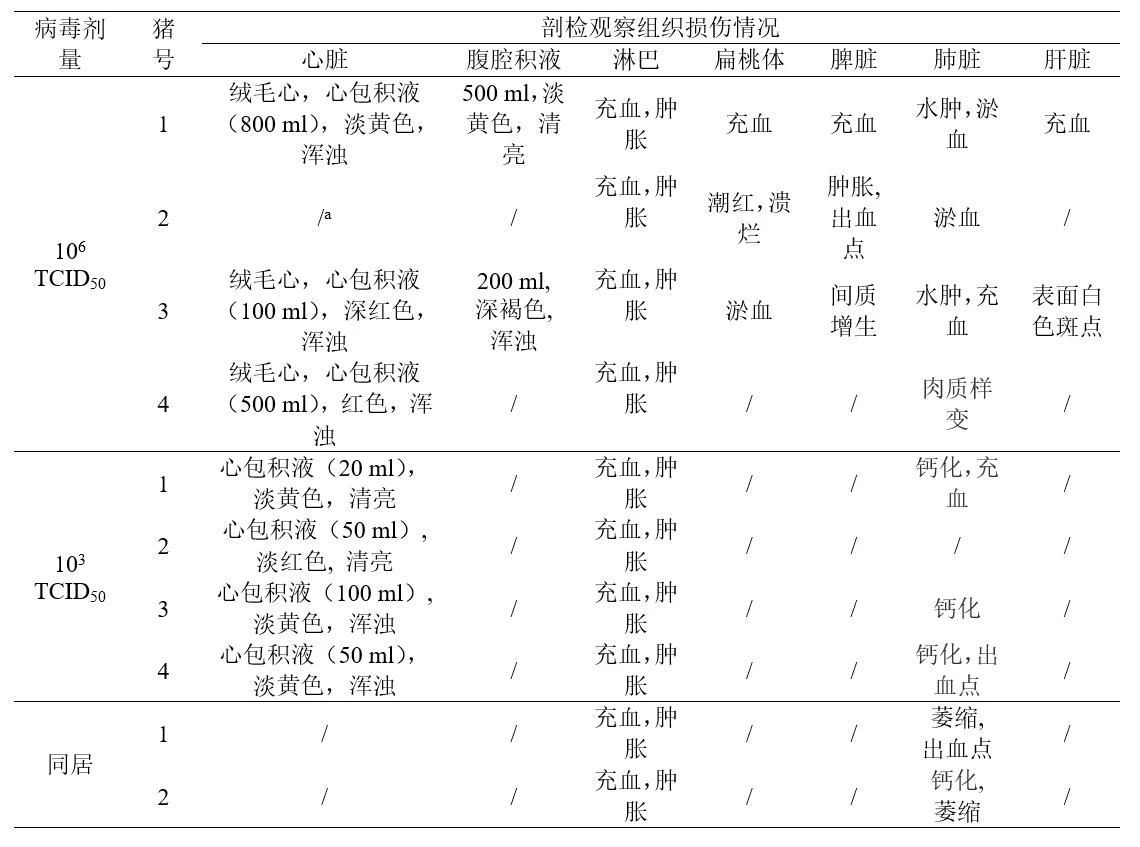
HeB/Q3/20 接种组的2头同居猪有1头猪在第8天从口腔拭子中检测出病毒核酸,第6天从肛拭子中检测出病毒核酸;第17天在两头猪的血液都检测到病毒核酸;两头猪所出现的发病病程与症状与103 TCID50攻毒组猪相似(表2),其中1头同居猪第24天发病死亡,另一头28天观察期内存活(图4)。
ASFV已在我国流行两年有余,但有关流行毒株的进化以及致病性表型研究鲜有报道。本研究报告了2020年6月至12月在我国部分省份开展的较为系统的ASFV流行病监测工作,共分离出22株基因II型ASFV流行毒株。与最早的分离株HLJ/18相比,所有2020年分离株基因组序列均发生不同程度的改变,包括核苷酸突变、缺失、插入或短片段替换等。其中11株病毒的EP402R基因呈现四种不同类型的核苷酸自然突变或缺失,造成这些CD2v蛋白编码缺陷,失去HAD表型。选取4株代表性病毒进行了动物回归试验,发现具有HAD表型的2株病毒致病力与HLJ/18相当,表现为典型高致死性;而2株缺失HAD表型分离株显示出较低的毒力,但依然表现出明显的毒力残留,高剂量感染能引起亚急性和慢性病程,并部分致死感染猪只;低剂量接种主要引起非致死性的持续感染,以及亚急性或慢性病程。两株病毒均具有很强的水平传播能力,尤其HeB/Q3/20表现更为突出。
当前尚无批准疫苗用于ASF的防控。ASF防控主要依靠对感染动物的快速诊断和及时清除。本研究发现,田间至少存在4种以上的non-HAD低致死率基因II型自然变异株,虽然致病力较典型强毒株明显降低,但仍然呈现明显的残留毒力,且具有很强的水平传播能力,很可能已在田间猪群中广泛传播,造成持续的感染、慢性病程甚至死亡,给非洲猪瘟早期诊断带来巨大的障碍,为我国非洲猪瘟防控带来全新的挑战,必须加以高度重视,制定和采取应对策略。
中国农业科学院哈尔滨兽医研究所国家非洲猪瘟专业实验室孙恩成副研究员、张振江博士、王子龙博士、何希君研究员为论文共同第一作者,步志高研究员和赵东明研究员为共同通讯作者。该研究得到国家重点研发项目(2018YFC1200601)等项目资助。
原文链接:
http://engine.scichina.com/doi/10.1007/s11427-021-1904-4
新闻推荐

